|
|
组成结构
|
视觉系统
视觉系统是神经系统的一个组成部分,它使机体具有了视知觉能力。 它使用可见光信息构筑机体对周围世界的感知。视觉系统具有将外部世界的二维投射重构为三维世界的能力。除了人类外,不同物种也有视觉系统,所能感知的可见光处于光谱中的不同位置。例如,有些物种可以看到紫外部分,而另一些则可以看到红外部分。
听觉系统
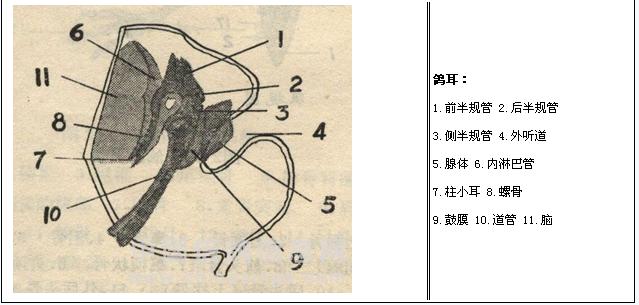
听觉系统是听觉的感觉系统,听觉指的是声源振动引起空气产生疏密波(声波),通过外耳和中耳组成的传音系统传递到内耳,经内耳的环能作用将声波的机械能转变为听觉神经上的神经冲动,后者传送到大脑皮层听觉中枢而产生的主观感觉。声波是由于四周的空气压力有节奏的变化而产生,当对象在震动时,四周的空气也会被影响。当物件越近,空气的粒子会被压缩;当物件越远,空气的粒子会被拉开。
体感系统
体感系统是有触觉、压觉、温觉、痛觉和本体感觉(关于肌肉和关节位置和运动、躯体姿势和运动以及面部表情的感觉)的一个系统。体感是和特殊感觉相对的一个概念。这些不同的体感模式源自不同类型的感受器。在哺乳类,体感的上行神经通路起源自身体不同部位的感受器,途经后柱-内侧丘系通路、脊髓丘脑前束、脊髓皮层前束和脊髓皮质后束,终于大脑皮层后中央回的体感皮层。
味觉系统
味觉系统是指感受味觉的感受器。对于哺乳类动物,其味觉系统是由口腔内的舌头以及连接舌头及大脑之间的神经系统组成。味觉系统的作用,主要是作为一个防卫机制,减少进食有问题的食物的机会。
嗅觉系统
嗅觉系统是指感受嗅觉的感受器。大部份哺乳类及爬虫类动物的嗅觉系统由主要嗅觉系统(main olfactory system)及辅助嗅觉系统(accessory olfactory system)组成,前者负责感应气态物质的气味,后者则负责感应液态物质的气味。 |
|
|
感觉系统的原理
|
外界刺激经过感受器(如眼的视网膜、内耳的柯蒂氏器等)时,触发的神经冲动由感觉神经传入中枢神经系统,再分别经特异投射系统和非特异投射系统达到大脑皮质,进而在大脑皮质发生加工,产生感觉。
(一)特异投射系统和非特异投射系统
感受器发出的神经冲动通过两条作用不同的神经通路传入大脑皮质,一条是特异性投射系统,另一条是非特异性投射系统。
1.特异性投射系统
特异性投射系统(specific projection system)是指各类感受器以特异的传入路径将神经冲动传送至大脑皮质的特定投射区的一种传入系统,如传导本体感觉冲动的薄束和楔束、传导体表感觉冲动的脊髓丘脑束以及听觉传导路、视觉传导路等。特异投射系统主要包括视觉、听觉、味觉、嗅觉、肤觉和本体感觉等。它们的作用是引起特定的感觉。
在特异投射系统中,丘脑是一个重要的转换站。除嗅觉外,所有的感觉纤维在到达大脑皮质之前都终止于丘脑,在此更换神经元后,发出的纤维将各种感觉冲动分别投射到大脑皮质的特定区域。丘脑对感觉信息进行初步的分析、综合。但它不能确定感觉的性质和强度,是皮质下较高级的感觉中枢。丘脑病变,往往引起感觉异常,如感觉过敏或感觉减退等。
2.非特异性投射系统
非特异性投射系统(nonspecific projection system)是指特异性传入系统神经束经过脑干时,发出侧支,与脑干网状结构内的神经元建立突触联系,然后经丘脑内侧部弥散性地投射到大脑皮质的广泛区域,不产生特定感觉的一种传入系统。网状结构系统是保证调节皮质紧张度和觉醒状态的脑器官。网状结构由许多中间神经元组成,它们与大脑皮质、下丘脑、脊髓、小脑的神经细胞有着广泛的神经联系。网状结构系统分上行网状系统和下行网状系统。上行网状系统对皮质的激活和紧张度的调节起着决定性的作用。实验证明,刺激除嗅觉以外的感受器都可以在网状结构的某些神经元得到冲动发放。这些冲动发放可以引起动物的觉醒反应。破坏网状结构,动物长期处于昏睡状态。因此,正常情况下人的各种心理活动都必须有网状激活系统的参加,因为人的绝大多数心理活动都是在清醒状态下进行的。下行网状系统则对效应器官的活动具有激活和抑制作用,它是大脑皮质对行为、活动控制必不可少的脑器官。 |
|
|
大脑皮质的感觉代表区
|
特异投射系统将来自感受器的冲动传向大脑皮质的特定区域,从而引起特定的感觉。现已探明大脑皮质的感觉代表区有以下几种。
1.体表感觉代表区
体表感觉主要指皮肤上的触、冷、温、痛等感觉。其大脑皮质代表区在中央后回(见图2――4)。这一区域的感觉投射有如下特点。(1)感觉传入的皮质投射是交叉的,即一侧的躯体感觉投射到对侧的大脑皮质的相应代表区。(2)感觉传入的皮质投射是倒置的,即下肢的感觉投射于这一区域的顶部,上肢的感觉投射于这一区域的中部,头颈部的感觉投射于这一区域的下部。(3)大脑皮质代表区的大小与身体不同部位的感觉灵敏度有关。感觉灵敏的部位所占的代表区域较大,如手、唇、口腔感觉的代表区域就特别大;感觉不灵敏的部位,如躯体的代表区就很小。这种结构特点有利于人进行精细的感觉分析,它是人类长期进化的结果。
2.本体感觉代表区
本体感觉指肌肉、关节的运动和位置感觉。中央前回是运动区,也是本体感觉的投射区。刺激中央前回也会引致病人企图发动肢体运动的主观感觉。
3.视觉代表区
视觉代表区在枕叶距状裂两侧。刺激该区域,可以使患者产生简单的主观光感觉,但不能引起完善的视觉形象。因为起源于鼻侧视网膜的传入纤维在视交叉处越至对侧,实行交叉,投射到对侧枕叶,而起源于颞侧的传入纤维并不交叉,直接投射到同侧枕叶。这样,一侧枕叶皮质主要与两眼同侧的视网膜相联系,因而与两眼的对侧视野有关,即右侧枕叶主要与两眼的左侧视野有关,而左侧枕叶主要与两眼的右侧视野有关。临床实践证明,一侧枕叶皮质受损害造成对侧偏盲,双侧枕叶皮质受损害造成全盲。
4.听觉代表区
听觉代表区在颞叶的颞横回。电刺激该区可以使患者产生铃声样或风吹样的主观音觉。听觉冲动的投射是双侧性的,即一侧皮质代表区与两侧耳的感受器都有关。因此,一侧颞叶皮质受损害并不影响听觉,只有左右两侧听觉代表区同时受损害,才产生完全的耳聋。
5.嗅觉和味觉代表区
尚未取得一致意见。一般认为,嗅觉冲动主要投射于海马回沟和海马回前部一带,味觉冲动投射于中央后回的头面部感觉投射区的下侧。
6.内脏感觉代表区
一般认为内脏感觉代表区在边缘叶。大脑半球内侧面皮质与间脑交接处的边缘及胼胝体旁的环周结构,称为边缘叶。它与附近的皮质(额叶眶部、岛叶、颞极、海马及齿状回等)以及有关的皮质下结构(包括隔区、杏仁核、丘脑及中脑被盖等),在结构与功能上相互间有密切的联系,构成一个统一的功能系统,称为边缘系统。边缘系统不仅与内脏有关,还与嗅觉、情绪、记忆等心理活动有关。 |
|
|
感觉器官
|
也叫感受器,是指分布在体表或组织内部的一些专门感受机体内、外环境改变的结构或装置。感受器的组成形成是多种多样的:有些感受器就是外周感觉神经末梢本身,如体表或组织内部与痛觉感受有关的游离神经末梢;有的感受器是裸露在神经末梢周围再包绕一些特殊的、由结缔组织构成的被膜样结构;但是对于一些与机体生存密切相关的感觉来说,体内存在着一些结构和功能上都高度分化了的感受细胞,它们以类似突触的形式直接或单位同感觉神经末梢相联系,如视网膜中的视杆和视锥细胞是光感受细胞,耳蜗中的毛细胞是声波感受细胞等,这些感受细胞连同它们的非神经性附属结构,构成了各种复杂的感觉器官如眼、耳等。高等动物中最重要的感觉器官,如眼、耳、前庭、嗅、味等器官,都分布在头部,称为特殊感官。
感受器官适宜刺激
各种感受器的一个共同功能特点,是它们各有自己最敏感、最容易接受的刺激形式;这就是说,用某种能量形式的刺激作用于某种感受器时,只需要极小的强度(即感觉阈值)就能引起相应的感觉。这一刺激形式或种类,就称为该感受器的适宜刺激,如在一定波长的电磁波是视网膜光感受细胞的适宜刺激,一定频率的机械震动是蜗毛细胞的适应刺激等。正因为如此,机体内、外环境中所发生的各种形式的变化,总是先作用于和它们相对应的那种感受器。这一现象的存在,是因为动物在长期的进化过程中逐步形成了具有各种特殊结构和功能 的感受器以及相应的附属结构的结果,便得它们有可能对内、外环境中某些有意义的变化进入灵敏的感受和精确的分析。不同动物所处的生活环境和条件不同,因此在进化中有可能形成一些异于人体的特殊感受装置,这在广大的动物界屡见不鲜,早已引起人们极大的兴趣和注意。研究这些可能是极低等动物的特殊感受装置,不仅对理解感受器活动的一般规律有帮助,而且有很大的仿生学意义。
感受器的换能作用
各种感受器在功能上的另一个共同特点,是能把作用于它们的各种刺激形式,转变成为相应的传入神经末梢或感受细胞的电反应,前者称为发生器电位(generator potential),在后者称为感受器电位(receptor potential)。如在第二章所述,发生器电位和感受器电位的出现,实际上是传入纤维的膜或感受细胞的膜进行了跨膜信号传递或转换过程的结果。和体内一般细胞一样,所有感受器细胞对外来不同刺激信号的跨膜转换,也主要是通过两种基本方式进行的,如声波振动的感受与蜗毛顶部膜中与听毛受力有关的机械细胞对外来中与听毛受力有关的机械门控通道的开放和关闭有关,这使毛细胞出现与声波振动相一致的感受器电位(即微音器电位);视杆和视锥细胞则是由于它们的外段结构中视盘膜上存在有受体蛋白(如视紫红质),它们在吸收光子后,再通过特殊的G-蛋白和作为效应器酶的磷酸二酯酶的作用,引起光感受器细胞外段胞浆中cGMP的分解,最后使外段膜出现感受器电位。在其他一些研究过的感受器,也看到了类似的两种信号转换机制。由此可见,所有感受性神经末梢 和感受器细胞出现电位变化,就是通过跨膜信号转换,把不同能量形式的外界刺激都转换成跨膜电位化的结果。
如前,发生器电位和感受器电位同终板电位和突触后电位一样,是一种过渡性慢电位,它们不具有“全或无”的特性而其幅度与外界刺激强度成比例;它不能作远距离传播而可能在局部实现时间性总和和空间性总和。正因为如此,感受器电位和发生器电位的幅度、持续时间和波动方向,就反映了外界刺激的某些特征,也就是说,外界刺激信号所携带的信息,也在换能过程中转移到了这种过渡性电变化的可变动的参数之中。
发生器电位和感受器电位的产生并不意味着感受器功能作用的完成,只有当这些过渡性电变化最终触发分布在该感受器的传入神经纤维上产生“全或无”式的可作远距离传导的动作电位序列时,才标志着这一感受器或感觉器官作用的完成;但主观感受的产生、对外界刺激信号的精细分析以及最后引起整个机体出现应答性反应和信息贮存等过程,则是传入神经纤维所输的神经信号到达各级脑中枢以后的反应,不属本章的内容。
感受器的编码作用
感受器在把外界刺激转换成神经动作电位时,不仅仅是发生了能量形式的转换;更重要的是把刺激所包涵的环境变化的信息,也转移到了新的电信号系统即动作电位的序列之中,即编码作用。编码(encoding)一词,本是工程通讯理论中的一个概念,指一种信号系统(如莫尔斯电码)如何把一定的信息内容(如电文内容)包涵在少量特定信号的排列组合之中。因此,感受器将外界刺激转变成神经动作电位的序列时,同时也实现了编码作用;中枢就是根据这些电信号序列才获得对外在世界的认识的。问题是外界刺激的质和量以及其他属性,是如何编码在特有的电信号序列中的?这一问题十分复杂,还远远没有弄清楚,下面先从最简单的方面加以叙述。
首先考虑外界刺激的“ |